Max-Planck-Institut für Multidisziplinäre Naturwissenschaften
Vielteilchenphysik erklärt Zelladhäsion
Forschungsbericht
28. September 2021
Wie das kooperative und kollektive Verhalten zwischen Zellen während der Zelladhäsion unter der Wirkung externer, mechanischer Kräfte entsteht, erklären Kristian Blom und Aljaž Godec in ihrer neuesten Publikation. Die Wissenschaftler am Göttinger Max-Planck-Institut für biophysikalische Chemie liefern damit einen mathematischen Beweis für die Existenz eines neuartigen dynamischen Phasenübergangs. Außerdem diskutieren sie, warum dieses Zellverhalten für den Gewebeumbau, Krebsmetastasen und Immunreaktionen relevant ist. (Physical Review X, 27. September 2021)
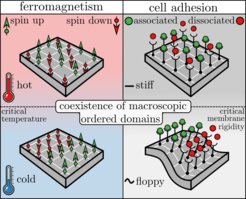
Analogie zwischen Ferromagnetismus und Zelladhäsion. Linke Spalte: 16 Spins in ferromagnetischem Material. Bei hohen Temperaturen dominiert die Entropie, so dass die Spins im gesamten Gitter zufällig nach oben oder unten ausgerichtet sind. Bei niedrigen Temperaturen, genauer gesagt unterhalb der Curie-Temperatur, dominiert die Energie, was zu makroskopisch geordneten Bereichen führt, in denen die Spins in dieselbe Richtung (nach oben oder unten) zeigen. Rechte Spalte: Ein Stück einer Zellmembran mit 16 Adhäsionsbindungen. Im Falle einer steifen Membran dominiert die Entropie, so dass die Bindungen im gesamten Gitter zufällig assoziiert oder dissoziiert sind. Bei einer geringeren Membransteifigkeit, genauer gesagt unterhalb der thermodynamisch kritischen Membransteifigkeit, koexistieren makroskopische Bereiche mit nur assoziierten bzw. dissoziierten Bindungen, die sich durch einen als thermodynamischen Phasenübergang bezeichneten Prozess bilden.
Analogie zwischen Ferromagnetismus und Zelladhäsion. Linke Spalte: 16 Spins in ferromagnetischem Material. Bei hohen Temperaturen dominiert die Entropie, so dass die Spins im gesamten Gitter zufällig nach oben oder unten ausgerichtet sind. Bei niedrigen Temperaturen, genauer gesagt unterhalb der Curie-Temperatur, dominiert die Energie, was zu makroskopisch geordneten Bereichen führt, in denen die Spins in dieselbe Richtung (nach oben oder unten) zeigen. Rechte Spalte: Ein Stück einer Zellmembran mit 16 Adhäsionsbindungen. Im Falle einer steifen Membran dominiert die Entropie, so dass die Bindungen im gesamten Gitter zufällig assoziiert oder dissoziiert sind. Bei einer geringeren Membransteifigkeit, genauer gesagt unterhalb der thermodynamisch kritischen Membransteifigkeit, koexistieren makroskopische Bereiche mit nur assoziierten bzw. dissoziierten Bindungen, die sich durch einen als thermodynamischen Phasenübergang bezeichneten Prozess bilden.
Wenn sich Zellen aneinander oder an eine extrazelluläre Matrix binden („kleben“), spricht man von „Zelladhäsion“. Dieses Phänomen ist von enormer Bedeutung für diverse biologische Prozesse. Ursprünglich wurde angenommen, dass die Biochemie der Adhäsionsbindungen bestimmt, wie stark sich die Zellen binden. Tatsächlich scheint es aber die Mechanik, insbesondere die Steifigkeit der zellulären Membran zu sein, die die Bindungskraft entscheidend beeinflusst, wenn nicht sogar dominiert. Auch wie Wechselwirkungen zwischen benachbarten Adhäsionsbindungen durch thermische Schwankungen der zellulären Membran zustande kommen, ist mittlerweile gut erforscht. Im Gegensatz dazu bleibt weiter unklar, inwieweit diese Wechselwirkungen die Stärke und Kinetik der Adhäsion regulieren. Weiter ist nicht bekannt, ob die Wechselwirkungen beeinflussen, wie die Bindungen auf die mechanischen Kräfte reagieren, die in physiologischen und pathologischen Prozessen auftreten.
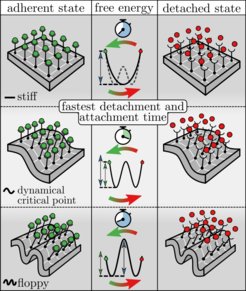
Ablöse- und Anhaftungszeiten sowie der dynamisch kritische Punkt. Äußere Spalten: Ein Stück einer Zellmembran mit 16 Adhäsionsbindungen, die alle assoziiert (links) oder dissoziiert (rechts) sind. Von oben nach unten: eine steife (stiff) Membran (oberhalb der thermodynamisch kritischen Membransteifigkeit); dynamisch kritische Membransteifigkeit; eine schlaffe (floppy) Membran (unterhalb der dynamisch kritischen Membransteifigkeit). In der mittleren Spalte sind die entsprechenden freien Energielandschaften entlang des Anteils der dissoziierten Bindungen dargestellt, wobei die beiden Endpunkte den vollständig assoziierten (grüner Kreis) und den vollständig dissoziierten (roter Kreis) Zustand widerspiegeln. Unterhalb der dynamisch kritischen Membransteifigkeit ist der Übergang zu einem vollständig assoziierten/dissoziierten Zustand (und damit die linke/rechte freie Energiebarriere) immer geschwindigkeitsbegrenzend, während die detaillierte Form der freien Energielandschaft von der Membransteifigkeit abhängt. Wenn die Steifigkeit größer als die thermodynamisch kritische Membransteifigkeit ist, hat die Landschaft eine einzige Vertiefung, während sie zwei Minima aufweist, wenn die Steifigkeit kleiner als die thermodynamisch kritische Membransteifigkeit ist. Letzteres entspricht einer Koexistenz von dichten und verdünnten makroskopischen Bereichen assoziierter Bindungen. Am dynamisch kritischen Punkt (Mitte) ist die Barriere zum vollständig assoziierten bzw. dissoziierten Zustand am kleinsten, was zur schnellsten Bindungs-/Lösungsrate führt. Jenseits dieses Punktes wird der thermodynamische Phasenübergang geschwindigkeitsbegrenzend, während die vollständig assoziierten und dissoziierten Zustände daraufhin durch typische Dichtefluktuationen erreicht werden.
Ablöse- und Anhaftungszeiten sowie der dynamisch kritische Punkt. Äußere Spalten: Ein Stück einer Zellmembran mit 16 Adhäsionsbindungen, die alle assoziiert (links) oder dissoziiert (rechts) sind. Von oben nach unten: eine steife (stiff) Membran (oberhalb der thermodynamisch kritischen Membransteifigkeit); dynamisch kritische Membransteifigkeit; eine schlaffe (floppy) Membran (unterhalb der dynamisch kritischen Membransteifigkeit). In der mittleren Spalte sind die entsprechenden freien Energielandschaften entlang des Anteils der dissoziierten Bindungen dargestellt, wobei die beiden Endpunkte den vollständig assoziierten (grüner Kreis) und den vollständig dissoziierten (roter Kreis) Zustand widerspiegeln. Unterhalb der dynamisch kritischen Membransteifigkeit ist der Übergang zu einem vollständig assoziierten/dissoziierten Zustand (und damit die linke/rechte freie Energiebarriere) immer geschwindigkeitsbegrenzend, während die detaillierte Form der freien Energielandschaft von der Membransteifigkeit abhängt. Wenn die Steifigkeit größer als die thermodynamisch kritische Membransteifigkeit ist, hat die Landschaft eine einzige Vertiefung, während sie zwei Minima aufweist, wenn die Steifigkeit kleiner als die thermodynamisch kritische Membransteifigkeit ist. Letzteres entspricht einer Koexistenz von dichten und verdünnten makroskopischen Bereichen assoziierter Bindungen. Am dynamisch kritischen Punkt (Mitte) ist die Barriere zum vollständig assoziierten bzw. dissoziierten Zustand am kleinsten, was zur schnellsten Bindungs-/Lösungsrate führt. Jenseits dieses Punktes wird der thermodynamische Phasenübergang geschwindigkeitsbegrenzend, während die vollständig assoziierten und dissoziierten Zustände daraufhin durch typische Dichtefluktuationen erreicht werden.
Das haben Blom und Godec nun untersucht. Für ihre Studie haben die Göttinger Forscher den Adhäsionsprozess auf einem zweidimensionalen, dynamischen Spin-Modell mit folgenden Einflussfaktoren abgebildet: wechselwirkende Adhäsionsbindungen mit einer intrinsischen Affinität und unter Einwirken einer externen Kraft, die zu jedem Zeitpunkt auf alle assoziierten Adhäsionsbindungen gleichmäßig verteilt wird. Die Wissenschaftler liefern einen mathematischen Beweis für die Existenz eines neuartigen dynamischen Phasenübergangs, bei dem Adhäsionsdomänen am schnellsten entstehen (beziehungsweise sich auflösen) und sich die kollektive Vielteilchendynamik qualitativ verändert. Variationen in der Steifigkeit der zellulären Membran und von externen Kräften beeinflussen die Adhäsionsbindungen am stärksten in der Nähe des entsprechenden dynamischen „kritischen Punktes“.
Die Arbeit bietet neue Einblicke in die statistische Physik von Vielteilchensystemen und schlägt eine Brücke von abstrakter Physik zu zellulärer und molekularer Biologie. In Zukunft wollen Blom und Godec ein noch detaillierteres theoretisches Modell der Zelladhäsion erstellen, das Bindungen einbezieht, die sich unter Kraft verstärken – so genannte „catch-bonds“. Damit möchten sie ein tieferes Verständnis der Zelladhäsion unter Krafteinwirkung erreichen. (kb/ag)
Ein Team um Bert de Groot hat jetzt mithilfe von Molekulardynamik-Simulationen erstmals in atomarer Auflösung sichtbar gemacht, wie Kaliumionen einen Kaliumionenkanal passieren. Die Simulationen könnten dazu beitragen, besser zu verstehen, wie Ionenkanäle reguliert werden oder Medikamente auf Ionenkanäle wirken.
Die im Januar 2025 gestartete Ausgründung des Max-Planck-Instituts für Multidisziplinäre Naturwissenschaften entwickelt neuartige Therapeutika, um die Qualität menschlicher Eizellen zu verbessern. Die Wirkstoffe sollen Frauen dabei helfen, länger fruchtbar zu bleiben und die Chancen auf eine erfolgreiche Schwangerschaft sowie auf eine In-vitro-Fertilisation (IVF) erhöhen.
Forschende um Victor Macarrón-Palacios, Elisa D‘Este und Manfred Kilimann am MPI für medizinische Forschung in Heidelberg und unserem Institut haben nun herausgefunden, dass das Protein Paralemmin-1 eine Schlüsselrolle für die Organisation des membranassoziierten periodischen Skeletts in Nervenzellen spielt.
Forschende um Peter Lenart und Monica Gobran haben eine bisher unbekannte Funktion der Proteinkinase Plk1 entdeckt: Sie steuert, dass Zellen rechtzeitig beginnen, sich zu teilen. Wird das Protein gehemmt, teilen sich Zellen erst viele Stunden später. Die neuen Ergebnisse könnten zu einem verbesserten Ansatz für Krebstherapien beitragen.
Forschende um Hauke Hillen konnten erstmals zeigen, wie sich das Erbgut des Nipah-Virus in infizierten Zellen vervielfältigt. Der Erreger kann beim Menschen eine tödlich verlaufende Gehirnentzündung auslösen. Mittels Kryo-Elektronenmikroskopie gelang es dem Team, die dreidimensionale Struktur der viralen „Kopiermaschine“ sichtbar zu machen.
Bereits vor der Geburt einer Frau sind ihre Eizellen in ihrem Körper angelegt. Mit zunehmendem Alter häufen sich jedoch DNA-Schäden in den Zellen an. Forschende haben jetzt aufgedeckt, warum die zelleigenen Reparaturmechanismen diese Schäden nicht beseitigen.
Göttinger Forschende haben erstmal eine molekulare „Feder“ entdeckt, die Ionenkanäle in den Hörsinneszellen im Ohr öffnen. Ihre Ergebnisse werfen neues Licht auf den Hörsinn und die Funktion von Ionenkanälen.
Forscher um Stefan Hell haben die Auflösung in der Fluoreszenzmikroskopie um das 30-fache gesteigert, ohne Fluoreszenzmoleküle nacheinander in einen leuchtenden und dunklen Zustand zu schalten. Indem sie einen Beleuchtungsstrahl mit einem Intensitätsminimum – einen dunklen Streifen im Strahl – verwendeten, konnten sie Moleküle bis auf einige wenige Nanometer trennen.
Forschenden ist es zum ersten Mal gelungen, den gesamten Prozess des Eisprungs in Follikeln einer Maus in Echtzeit sichtbar zu machen. Ihre neue Lebendzellmikroskopie-Methode ermöglicht es, den Eisprung mit hoher räumlicher und zeitlicher Auflösung zu untersuchen und eröffnet neue Möglichkeiten in der Fruchtbarkeitsforschung.
Einem Team vom Göttinger MPI für Multidisziplinäre Naturwissenschaften und vom Heidelberger MPI für medizinische Forschung ist es gelungen, mit der MINFLUX-Mikroskopie-Methode Abstände innerhalb von Biomolekülen bis hinunter zu 1 Nanometer zu messen, und zwar mit Ångström-Präzision.
Alzheimer ist die häufigste Ursache von Demenz. Forschende zeigten jetzt: Nicht nur Nervenzellen produzieren das Protein Beta-Amyloid, das im Gehirn von Erkrankten zu schädlichen Plaques verklumpt und Nervenzellen schädigt. Auch Gliazellen bilden das Protein und tragen zur Plaquebildung bei.
Kleine biologische Systeme sind aufgrund von thermischen Fluktuationen von Natur aus stochastisch. Die Inferenz der Dissipation, d. h. ob und inwieweit biologische Systeme (oder Prozesse) aus dem Gleichgewicht gebracht werden, aus fluktuierenden Beobachtungen, die in der Regel nur eine kleine Teilmenge von Freiheitsgraden erfassen, bleibt eine zentrale Herausforderung in der modernen Nichtgleichgewichtsphysik.
 zeigen. Rechte Spalte: Ein Stück einer Zellmembran mit 16 Adhäsionsbindungen. Im Falle einer steifen Membran dominiert die Entropie, so dass die Bindungen im gesamten Gitter zufällig assoziiert oder dissoziiert sind. Bei einer geringeren Membransteifigkeit, genauer gesagt unterhalb der thermodynamisch kritischen Membransteifigkeit, koexistieren makroskopische Bereiche mit nur assoziierten bzw. dissoziierten Bindungen, die sich durch einen als thermodynamischen Phasenübergang bezeichneten Prozess bilden.")
 oder dissoziiert (rechts) sind. Von oben nach unten: eine steife (stiff) Membran (oberhalb der thermodynamisch kritischen Membransteifigkeit); dynamisch kritische Membransteifigkeit; eine schlaffe (floppy) Membran (unterhalb der dynamisch kritischen Membransteifigkeit). In der mittleren Spalte sind die entsprechenden freien Energielandschaften entlang des Anteils der dissoziierten Bindungen dargestellt, wobei die beiden Endpunkte den vollständig assoziierten (grüner Kreis) und den vollständig dissoziierten (roter Kreis) Zustand widerspiegeln. Unterhalb der dynamisch kritischen Membransteifigkeit ist der Übergang zu einem vollständig assoziierten/dissoziierten Zustand (und damit die linke/rechte freie Energiebarriere) immer geschwindigkeitsbegrenzend, während die detaillierte Form der freien Energielandschaft von der Membransteifigkeit abhängt. Wenn die Steifigkeit größer als die thermodynamisch kritische Membransteifigkeit ist, hat die Landschaft eine einzige Vertiefung, während sie zwei Minima aufweist, wenn die Steifigkeit kleiner als die thermodynamisch kritische Membransteifigkeit ist. Letzteres entspricht einer Koexistenz von dichten und verdünnten makroskopischen Bereichen assoziierter Bindungen. Am dynamisch kritischen Punkt (Mitte) ist die Barriere zum vollständig assoziierten bzw. dissoziierten Zustand am kleinsten, was zur schnellsten Bindungs-/Lösungsrate führt. Jenseits dieses Punktes wird der thermodynamische Phasenübergang geschwindigkeitsbegrenzend, während die vollständig assoziierten und dissoziierten Zustände daraufhin durch typische Dichtefluktuationen erreicht werden.")