Max-Planck-Institut für Multidisziplinäre Naturwissenschaften
Logistik auf kleinstem Raum: Transportprozesse zwischen Zellkern und Zytoplasma
Logistics on smallest possible space: Transport processes between cell nucleus and cytoplasm
Forschungsbericht (importiert) 2008 - Max-Planck-Institut für Multidisziplinäre Naturwissenschaften
Autoren
Dirk Görlich; Steffen Frey
Abteilungen
Zelluläre Logistik (Prof. Dr. Dirk Görlich) MPI für biophysikalische Chemie, Göttingen
Zusammenfassung
Der Zellkern verfügt über keine eigene Proteinsynthese, sondern importiert alle benötigten Proteine aus dem Zytosol. Umgekehrt versorgt er das Zytosol mit Ribosomen, mRNAs und tRNAs. Sämtlicher Kern-Zytoplasma-Transport wird durch die Permeabilitätsbarriere der Kernporen kontrolliert. Diese Permeabilitätsbarriere ist ein „intelligentes“ Hydrogel mit erstaunlichen Materialeigenschaften. Es unterdrückt den Durchtritt von inerten Makromolekülen, erlaubt aber einen bis zu 20.000fach schnelleren Einstrom derselben Moleküle, wenn diese an einen passenden Kern-Transport-Rezeptor gebunden sind.
Zusammenfassung
The cell nucleus is enclosed by the nuclear envelope, lacks protein synthesis and therefore imports each and every protein from the cytosol. Conversely, the nucleus supplies the cytoplasm with nuclear products, such as ribosomes, tRNAs and mRNAs. The permeability barrier of nuclear pore complexes controls all this exchange. This permeability barrier is an "intelligent" hydrogel with truly remarkable properties. It excludes inert macromolecules, but permits an up to 20 000-fold faster entry of cargoes, when these are bound to appropriate nuclear transport receptors.
Kompartimentierung
Die uns bekannten Lebewesen lassen sich in zwei Kategorien unterteilen: Prokaryoten, das heißt Bakterien oder Archaebakterien, sowie Eukaryoten, zu denen alle mit bloßem Auge sichtbaren Organismen wie Pflanzen, Tiere und Pilze, aber auch eine Reihe von Einzellern wie Hefen oder Amöben gehören. Das charakteristische Merkmal eukaryotischer Zellen ist deren Aufteilung in verschiedene Reaktionsräume, die Kompartimente genannt werden und Ausdruck einer strikt arbeitsteiligen Organisation sind. So enthält eine menschliche Zelle hunderte Mitochondrien, in denen die Zellatmung abläuft, und die für die Energieversorgung der Zelle zuständig sind. Sie besitzt zudem ein sekretorisches System, mit dessen Hilfe zum Beispiel Hormone produziert und aus der Zelle ausgeschleust werden. Und natürlich enthält sie den Zellkern, in dem die genetische Information aufbewahrt, verwaltet und abgerufen wird, und der gewissermaßen die Leitzentrale der Zelle darstellt. Die eben genannten Zellbestandteile „schwimmen“ in einer Grundsubstanz, die Zytosol genannt wird und ein ebenso eigenständiges Kompartiment darstellt.
Tierische oder pflanzliche Zellen haben also eine extrem komplizierte innere Struktur, deren Vorteile sich wahrscheinlich am eindrucksvollsten anhand der Tatsache zusammenfassen lassen, dass nur eukaryotische Organismen den Sprung zur Entwicklung komplexer, vielzelliger Lebewesen geschafft haben. Die Verwaltung des Genoms in einem dafür spezialisierten Kompartiment, nämlich dem Zellkern, hat sicher ganz entscheidend zu dieser Entwicklung beigetragen und erlaubt es Eukaryoten, eine rund 1.000fach größere Menge an genetischer Information zu handhaben als Bakterien. Sie können damit den „Bauplan“ eines hochkomplexen vielzelligen Organismus von Generation zu Generation weitergeben.
Kern-Zytoplasma-Transport ist essentiell für eukaryotisches Leben
Wie alles im Leben hat auch die ausgeklügelte strukturelle Organisation eukaryotischer Zellen ihren Preis und kann nur mit erheblichem Aufwand aufrechterhalten werden. Dieser Aufwand liegt nicht zuletzt in der Versorgung der einzelnen Kompartimente mit bestimmten Makromolekülen wie Enzymen, strukturellen Proteinen oder Ribonukleinsäuren. So werden Proteine fast ausschließlich im Zytosol synthetisiert und von dort auf Mitochondrien, den Zellkern oder andere Zellorganellen verteilt. Diese Verteilung erfolgt nicht zufällig; vielmehr wird beispielsweise ein Enzym, das der Zellkern benötigt, in aller Regel ausschließlich in den Kern importiert und eben nicht in die Mitochondrien.
Umgekehrt erfolgt auch ein massiver Export aus dem Kern. Wie bereits erwähnt, werden Proteine im Zytosol produziert. Die Baupläne für jedes einzelne Protein sind jedoch im Genom niedergeschrieben, das im Zellkern aufbewahrt wird und diesen auch nicht verlässt. Diese Information muss aber trotzdem aus dem Kern ins Zytoplasma gelangen. Die Lösung des Problems ist, dass Kopien einzelner Gene erstellt und aus dem Zellkern exportiert werden. Im Zytosol werden diese Kopien von den Ribosomen gelesen, die wiederum einzelne Aminosäuren, entsprechend der genetischen Vorschrift, zu Proteinen zusammenfügen. Auch Ribosomen werden im Zellkern produziert und müssen erst in das Zytoplasma exportiert werden.
Diese Import- und Exportprozesse zwischen Kern und Zytoplasma sind keineswegs seltene Ereignisse. Vielmehr erfordert jede einzelne enzymatische Reaktion im Kern, sei es das Kopieren von Genen oder die Reparatur von Schadstellen im Genom, Proteine, die zuvor im Zytoplasma produziert und in den Kern importiert wurden. Umgekehrt könnte kein einziges eukaryotisches Gen in ein Protein umgesetzt werden, ohne dass zuvor Ribosomen und Genkopien aus dem Kern exportiert worden wären.
Ein Blick in die Transportmaschinerie
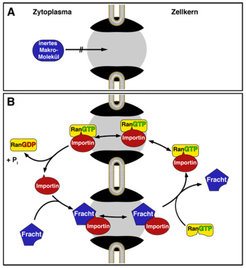
Abb. 1: Importin-abhängiger Proteintransport in den Zellkern. Zellkern und Zytoplasma sind durch die Kernhülle voneinander getrennt. Ein stofflicher Austausch zwischen beiden Kompartimenten kann daher nur durch Kernporen erfolgen. Die Permeabilitätsbarriere der Kernpore (grau) unterdrückt die Passage von „normalen“ (inerten) Makromolekülen (A), erlaubt jedoch eine praktisch ungehinderte Passage von Importinen und anderen Kerntransportrezeptoren (B). Importine binden ihre Fracht im Zytoplasma und entlassen sie, sobald sie auf RanGTP im Zellkern treffen. Der Importin-RanGTP-Komplex passiert die Permeabilitätsbarriere dann in entgegengesetzter Richtung, im Zytosol wird GTP hydrolysiert, Ran entlassen und das Importin kann erneut Fracht laden. Das Zusammenspiel mit dem RanGTPase-System macht das Importin zu einer Proteinpumpe. Die eigentliche Passage durch die Permeabilitätsbarriere ist jedoch ein reversibler und energieunabhängiger Prozess.
Abb. 1: Importin-abhängiger Proteintransport in den Zellkern. Zellkern und Zytoplasma sind durch die Kernhülle voneinander getrennt. Ein stofflicher Austausch zwischen beiden Kompartimenten kann daher nur durch Kernporen erfolgen. Die Permeabilitätsbarriere der Kernpore (grau) unterdrückt die Passage von „normalen“ (inerten) Makromolekülen (A), erlaubt jedoch eine praktisch ungehinderte Passage von Importinen und anderen Kerntransportrezeptoren (B). Importine binden ihre Fracht im Zytoplasma und entlassen sie, sobald sie auf RanGTP im Zellkern treffen. Der Importin-RanGTP-Komplex passiert die Permeabilitätsbarriere dann in entgegengesetzter Richtung, im Zytosol wird GTP hydrolysiert, Ran entlassen und das Importin kann erneut Fracht laden. Das Zusammenspiel mit dem RanGTPase-System macht das Importin zu einer Proteinpumpe. Die eigentliche Passage durch die Permeabilitätsbarriere ist jedoch ein reversibler und energieunabhängiger Prozess.
Der Zellkern ist von zwei Membranen umgeben, die für Proteine und andere Makromoleküle völlig undurchlässig sind. Der stoffliche Austausch kann daher nicht direkt durch diese Membranen erfolgen. Stattdessen sind in die Kernhülle so genannte Kernporen eingebettet, die man sich als hochselektive Tore vorstellen kann und die Teil einer ganzen Transportmaschinerie sind. Diese Transportmaschinerie ist das zentrale Forschungsthema der Abteilung „zelluläre Logistik“ am MPI für biophysikalische Chemie.
Importine und Exportine [1] bilden den mobilen Teil des Transportsystems (Abb. 1). Während die Kernporen für die meisten Makromoleküle ab einem bestimmten Größenlimit dicht verschlossen erscheinen, haben Importine und Exportine das Privileg, die Permeabilitätsbarriere der Kernporen nahezu ungehindert passieren zu können. Das Entscheidende dabei ist, dass sie bei ihrem Porendurchtritt auch „Fracht“ oder „Passagiere“ mitnehmen können. Nun darf nicht jeder Passagier „an Bord“, sondern Importine erkennen mit molekularer Präzision, welche Proteine in den Kern importiert werden sollen und welche nicht. Ein weiteres Problem ist, dass ein Importin nicht nur „seine“ Passagiere erkennen, sondern auch sicherstellen muss, dass die Fracht im Zytosol aufgenommen und im Kern entlassen wird. Um seine Fracht sicher an den Bestimmungsort bringen zu können, muss ein Importin erkennen, ob es sich innerhalb oder außerhalb des Kerns befindet. Das ist kein triviales Problem, und es stellte sich heraus, dass Importine dazu Instruktionen von einem molekularen Schalter, der „Ran“ genannt wird, erhalten. Dieser Schalter steht nur im Kern auf „an“ (RanGTP), und diese Schalterstellung gibt den Importinen das Zeichen, ihre Ladung zu löschen. Daraufhin kehrt das Importin unbeladen ins Zytosol zurück, um eine weitere Runde Import zu vermitteln.
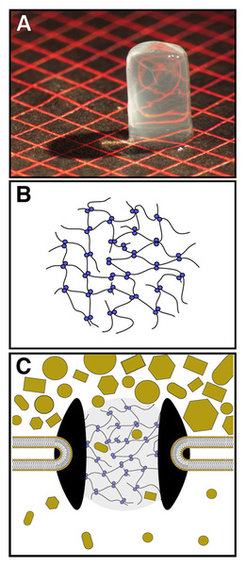
Abb. 2: Die Permeabilitätsbarriere der Kernpore – ein hochselektives Hydrogel. Die so genannten FG-Repeat-Domänen von Kernporenproteinen bestimmen die Permeabilitätseigenschaften von Kernporen. Ihre lokale Konzentration innerhalb der Kernpore ist mit geschätzten 100-200 mg/ml extrem hoch. Löst man die FG-Repeat-Domäne des Kernporenproteins Nsp1p zu einer so hohen Konzentration, so bildet sie ein transparentes, elastisches FG-Hydrogel, dessen mechanische Eigenschaften an Gummibärchen (ein Gelatine-Hydrogel) erinnern. Ein solches FG-Hydrogel wurde in A auf eine gemusterte Unterlage platziert und photographiert. FG-Repeats sind hydrophile, lineare Polymere, die in regelmäßigen Abständen hydrophobe Flecken (FG-Motive) enthalten. Die Hydrogel-Bildung beruht auf einer Quervernetzung zwischen einzelnen FG-Repeat-Domänen, die u. a. durch hydrophobe Interaktionen zwischen den FG-Motiven (blaue Punkte) vermittelt werden (siehe B). Das FG-Hydrogel kann daher als dreidimensionales Sieb betrachtet werden, das kleine Moleküle ungehindert passieren lässt, aber den Durchtritt von inerten Makromolekülen (ocker) unterdrückt (siehe C). Dabei bestimmt die Maschenweite die Ausschlussgrenze (≈ 5 nm Durchmesser).
Abb. 2: Die Permeabilitätsbarriere der Kernpore – ein hochselektives Hydrogel. Die so genannten FG-Repeat-Domänen von Kernporenproteinen bestimmen die Permeabilitätseigenschaften von Kernporen. Ihre lokale Konzentration innerhalb der Kernpore ist mit geschätzten 100-200 mg/ml extrem hoch. Löst man die FG-Repeat-Domäne des Kernporenproteins Nsp1p zu einer so hohen Konzentration, so bildet sie ein transparentes, elastisches FG-Hydrogel, dessen mechanische Eigenschaften an Gummibärchen (ein Gelatine-Hydrogel) erinnern. Ein solches FG-Hydrogel wurde in A auf eine gemusterte Unterlage platziert und photographiert. FG-Repeats sind hydrophile, lineare Polymere, die in regelmäßigen Abständen hydrophobe Flecken (FG-Motive) enthalten. Die Hydrogel-Bildung beruht auf einer Quervernetzung zwischen einzelnen FG-Repeat-Domänen, die u. a. durch hydrophobe Interaktionen zwischen den FG-Motiven (blaue Punkte) vermittelt werden (siehe B). Das FG-Hydrogel kann daher als dreidimensionales Sieb betrachtet werden, das kleine Moleküle ungehindert passieren lässt, aber den Durchtritt von inerten Makromolekülen (ocker) unterdrückt (siehe C). Dabei bestimmt die Maschenweite die Ausschlussgrenze (≈ 5 nm Durchmesser).
Im Gegensatz zu Importinen sind Exportine für den Transport von Ribonukleinsäuren und bestimmten Proteinen aus dem Zellkern zuständig. Sie funktionieren ganz analog zu den Importinen, nur interpretieren sie die Zeichen des molekularen Schalters Ran genau entgegengesetzt: Sie laden Fracht im Kern auf und entladen sie in das Zytosol.
Die eben beschriebenen Transportprozesse laufen in sämtlichen menschlichen Zellen ab, die typischerweise einen Durchmesser von einem hundertstel Millimeter haben. Sie sind damit ein Beispiel dafür, wie Logistik auf kleinstem Raum organisiert werden kann.
Offensichtlich spielen die Kernporen eine Schlüsselrolle in diesen Austauschprozessen. Sie repräsentieren eine Kategorie biologischer Objekte, die auch als molekulare Maschinen bezeichnet werden. Im Gegensatz zu Enzymen katalysieren diese nicht einfach nur chemische Reaktionen. Vielmehr bewerkstelligen molekulare Maschinen komplexere Aufgaben, wie beispielsweise das gezielte Bewegen von Objekten innerhalb einer Zelle oder eben das Sortieren von Proteinen in solche, die in den Kern gehören und solche, die im Zytoplasma verbleiben sollen.
Gemessen an der täglichen Erfahrung sind diese molekularen Maschinen winzig, und selbst mit den besten Lichtmikroskopen erscheint eine Kernpore nur als punktförmiges Objekt. In molekularem Maßstab sind Kernporen jedoch gigantisch: Sie setzen sich aus etwa 700 einzelnen Polypeptidketten bzw. 20 Millionen Einzelatomen zusammen. Entsprechend ihrer Größe sind Kernporen auch äußerst effektive Sortiermaschinen. Auf der einen Seite verhindern sie ein Vermischen der im Kern bzw. Zytosol gelösten Inhaltsstoffe und blockieren den Durchtritt vieler Makromoleküle. Gleichzeitig kann eine einzige Kernpore pro Sekunde aber bis zu 1.000 Importin-Moleküle mit gebundener Fracht selektiv passieren lassen, ohne dass die Präzision dieses Sortierprozesses darunter leidet [2]. Diese Transportkapazität ist aber auch unbedingt erforderlich, denn überschlägig gerechnet entspricht der Materialstrom, der täglich durch die rund 1016 Kernporen eines Menschen selektiv geleitet wird, in etwa unserem Körpergewicht.
Wie funktioniert die Permeabilitätsbarriere der Kernpore?
Derzeit beschäftigt uns ganz unmittelbar die Frage, warum Importine und Exportine nahezu ungehindert Kernporen passieren können, aber andere Objekte gleicher Größe mit hoher Präzision ausgefiltert werden. Eine entscheidende Rolle spielen dabei die so genannten Phenylalanin-reichen Repeat-Motive (FG-Repeats) [3], die Teil von Kernporenproteinen sind und in insgesamt ≈104 Kopien pro Kernpore vorkommen. Importine und Exportine binden FG-Repeats, und diese Interaktionen sind Voraussetzung für die erleichterte Translokation durch die Kernporen [4]. Wie aber derartige Interaktionen eine Beschleunigung der Kernporenpassage bewirken, ist keine triviale Frage, da eine bloße Bindung eines Importins oder Exportins an die FG-Repeats die Kernporenpassage lediglich verzögern sollte.
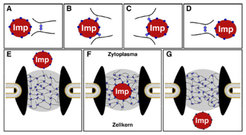
Abb. 3: Das Hydrogel-Modell der Kernporenfunktion. Die Permeabilitätsbarriere der Kernporen verhindert nicht nur ein unkontrolliertes Vermischen von Zytosol und Kerninhalt, sondern erlaubt auch sehr großen Objekten (bis zu einem Durchmesser von mehr als 25 nm), die Kernporen mithilfe eines Kern-Transportrezeptors (z.B. Importin) zu passieren. Die Größe solcher Transportkomplexe übersteigt die Maschenweite des FG-Hydrogels; sie müssen deshalb für ihren Durchtritt die im Wege liegenden Maschen des FG-Hydrogels kurzzeitig aufbrechen. Die Schemata A-D illustrieren diesen Prozess. Eine Masche wird durch Interaktion zwischen FG-Motiven (blau) gebildet und stabilisiert. Importine binden diese Motive, öffnen dadurch eine Masche und können das Hindernis samt ihrer Fracht passieren. Schemata E-F zeigen eine Kernpore mit FG-Hydrogel-Barriere. Importine (Imp) können Maschen lokal öffnen und sich im Hydrogel lösen. Sie werden dann gewissermaßen von der Permeabilitätsbarriere umflossen. Das stellt sicher, dass die Barriere intakt bleibt, selbst während große Objekte durch die Pore geschleust werden. Der im Gel gelöste Transportrezeptor kann die Barriere auf der anderen Seite wieder verlassen.
Abb. 3: Das Hydrogel-Modell der Kernporenfunktion. Die Permeabilitätsbarriere der Kernporen verhindert nicht nur ein unkontrolliertes Vermischen von Zytosol und Kerninhalt, sondern erlaubt auch sehr großen Objekten (bis zu einem Durchmesser von mehr als 25 nm), die Kernporen mithilfe eines Kern-Transportrezeptors (z.B. Importin) zu passieren. Die Größe solcher Transportkomplexe übersteigt die Maschenweite des FG-Hydrogels; sie müssen deshalb für ihren Durchtritt die im Wege liegenden Maschen des FG-Hydrogels kurzzeitig aufbrechen. Die Schemata A-D illustrieren diesen Prozess. Eine Masche wird durch Interaktion zwischen FG-Motiven (blau) gebildet und stabilisiert. Importine binden diese Motive, öffnen dadurch eine Masche und können das Hindernis samt ihrer Fracht passieren. Schemata E-F zeigen eine Kernpore mit FG-Hydrogel-Barriere. Importine (Imp) können Maschen lokal öffnen und sich im Hydrogel lösen. Sie werden dann gewissermaßen von der Permeabilitätsbarriere umflossen. Das stellt sicher, dass die Barriere intakt bleibt, selbst während große Objekte durch die Pore geschleust werden. Der im Gel gelöste Transportrezeptor kann die Barriere auf der anderen Seite wieder verlassen.
Zur Lösung dieses Paradoxons haben wir vorgeschlagen, dass die Bildung der Barriere und die erleichterte Kernporenpassage unmittelbar verknüpfte Phänomene darstellen [2; 5]. Da FG-Repeats für die erleichterte Translokation benötigt werden, sollten sie auch an der Bildung der Barriere beteiligt sein. FG-Repeats bestehen aus bis zu 50 kurzen Clustern hydrophober Aminosäuren (meist Phenylalanin, F), zwischen denen jeweils sehr hydrophile Abschnitte liegen. Durch hydrophobe Interaktion zwischen den hydrophoben Clustern sollte sich aus den zunächst linearen FG-Repeats ein dreidimensionales Netzwerk (d. h. ein reversibles Hydrogel) bilden (Abb. 2), das den Durchtritt solcher Objekte verhindert, deren Dimensionen die „Maschenweite“ übersteigt (Abb. 2C).
Importine und Exportine können jedoch direkt in das Netzwerk eingebettet werden, da sie selbst die hydrophoben Cluster der FG-Repeats binden und dadurch Inter-Repeat-Interaktionen lokal unterdrücken (Abb. 3). Gewissermaßen können sie sich also in der Permeabilitätsbarriere lösen und sie somit durchqueren.
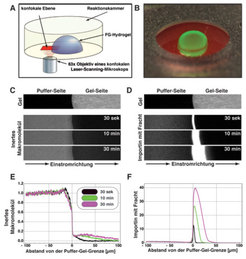
Abb. 4: Analyse der Permeabilitätseigenschaften des FG-Hydrogels. A). Schema des Messansatzes. Ein FG-Hydrogel wird so auf dem Objekttisch eines konfokalen Laser-Scanning-Mikroskops so positioniert, dass die Puffer/Gel-Grenze in der konfokalen Bildebene abgebildet wird. Der Einstrom zugegebener fluoreszenter Proteine in das Gel wird in Echtzeit mikroskopisch verfolgt. B) Aufnahme eines FG-Hydrogels, das mit einem inerten rot-fluoreszierenden Protein und einem grün-fluoreszierenden Importin-Fracht-Komplex inkubiert wurde. Das inerte Protein blieb ausgeschlossen, während der Importin-Komplex tief in das Gel eindrang. C)Durch konfokale Laser-Scanning-Mikroskopie wurden das far-red markierte FG-Hydrogel („Gel“) sowie der Einstrom eines rot-fluoreszierenden inerten Moleküls in dieses Gel visualisiert. Dieser Einstrom war sehr ineffizient, d.h. das Gel erwies sich als eine sehr effektive Barriere. D) Gleichzeitig wurde der Einstrom eines grün-fluoreszierenden Importin-Fracht-Komplexes verfolgt. Dieser Einstrom war so effizient, dass die Diffusion an die Gelgrenze zum geschwindigkeitsbestimmenden Schritt wurde. Der Komplex reicherte sich sehr stark im Gel an (Partitionskoeffizient > 100); die Aufnahmen sind daher im Gelbereich überexponiert. Entscheidend ist, dass der Importin-Fracht-Komplex nicht an der Puffer-Gel-Grenze stecken bleibt, sondern sich mit einer Geschwindigkeit in das Gel bewegt, die eine Kernporenpassage innerhalb von 10 Millisekunden gestatten würde. E) und F) Quantifizierungen der experimentellen Primärdaten von C und D. Dabei sind die Konzentrationsprofile der diffundierenden Moleküle zu verschiedenen Zeitpunkten ihres Einstroms in das FG-Hydrogel dargestellt.
Abb. 4: Analyse der Permeabilitätseigenschaften des FG-Hydrogels. A). Schema des Messansatzes. Ein FG-Hydrogel wird so auf dem Objekttisch eines konfokalen Laser-Scanning-Mikroskops so positioniert, dass die Puffer/Gel-Grenze in der konfokalen Bildebene abgebildet wird. Der Einstrom zugegebener fluoreszenter Proteine in das Gel wird in Echtzeit mikroskopisch verfolgt. B) Aufnahme eines FG-Hydrogels, das mit einem inerten rot-fluoreszierenden Protein und einem grün-fluoreszierenden Importin-Fracht-Komplex inkubiert wurde. Das inerte Protein blieb ausgeschlossen, während der Importin-Komplex tief in das Gel eindrang. C)Durch konfokale Laser-Scanning-Mikroskopie wurden das far-red markierte FG-Hydrogel („Gel“) sowie der Einstrom eines rot-fluoreszierenden inerten Moleküls in dieses Gel visualisiert. Dieser Einstrom war sehr ineffizient, d.h. das Gel erwies sich als eine sehr effektive Barriere. D) Gleichzeitig wurde der Einstrom eines grün-fluoreszierenden Importin-Fracht-Komplexes verfolgt. Dieser Einstrom war so effizient, dass die Diffusion an die Gelgrenze zum geschwindigkeitsbestimmenden Schritt wurde. Der Komplex reicherte sich sehr stark im Gel an (Partitionskoeffizient > 100); die Aufnahmen sind daher im Gelbereich überexponiert. Entscheidend ist, dass der Importin-Fracht-Komplex nicht an der Puffer-Gel-Grenze stecken bleibt, sondern sich mit einer Geschwindigkeit in das Gel bewegt, die eine Kernporenpassage innerhalb von 10 Millisekunden gestatten würde. E) und F) Quantifizierungen der experimentellen Primärdaten von C und D. Dabei sind die Konzentrationsprofile der diffundierenden Moleküle zu verschiedenen Zeitpunkten ihres Einstroms in das FG-Hydrogel dargestellt.
Kernporen sind extrem komplex aufgebaut. Um ihre Funktionsprinzipien wirklich begreifen zu können, muss das experimentelle System zunächst auf das Wesentliche reduziert werden. Wir konnten jetzt die Permeabilitätsbarriere im Reagenzglas aus einer einzigen FG-Repeat-Domäne eines Kernporenproteins nachbilden [6; 7]. Sie erwies sich in der Tat als ein Hydrogel; vergleichbar mit Götterspeise oder dem Glaskörper des Auges, die auch ein Hydrogel darstellen.
Im Gegensatz zu gewöhnlichen Hydrogelen hat „unser“ Hydrogel jedoch die phantastische Eigenschaft, "normalen" Makromolekülen ein Eindringen in das Gel zu verwehren (Abb. 4). Wird ein solches „normales“ Makromolekül jedoch mit einem Kern-Import-Signal versehen und an ein Importin gebunden, dann erhöht sich die Einströmgeschwindigkeit in das Gel bis zu 20.000fach, und das, obwohl die Bindung des Importins die Größe des diffundierenden Objektes auf ein Vielfaches erhöht.
Wir möchten jetzt verstehen, wie diese Prozesse im Detail ablaufen, das heißt welche Atomgruppen wie und wann mit anderen interagieren müssen, damit die Permeabilitätsbarriere die Selektivität erlangt, die Kernporen auszeichnet. Darüber hinaus möchten wir verstehen, wie Importine und Exportine ihre Fracht so zielsicher inmitten tausender anderer Proteinmoleküle erkennen und wie Kernporen aus ihren Vorstufen zusammengebaut und in die Kernhülle eingelassen werden.
Originalveröffentlichungen
1.
D. Görlich, U. Kutay:
Transport between the cell nucleus and the cytoplasm.
Annual Review of Cell and Developmental Biology 15, 607-660 (1999).
2.
K. Ribbeck, D. Görlich:
Kinetic analysis of translocation through nuclear pore complexes.
EMBO Journal 20, 1320-1330 (2001).
3.
M. Rout, S. Wente:
Pores for thought: nuclear pore complex proteins.
Trends in Cell Biology 4, 357-364 (1994).
4.
R. Bayliss, K. Ribbeck, D. Akin, H. M. Kent, C. M. Feldherr, D. Görlich, M. Stewart:
Interaction between NTF2 and xFxFG-containing nucleoporins is required to mediate nuclear import of RanGDP.
Journal of Molecular Biology 293, 579-593 (1999).
5.
K. Ribbeck, D. Görlich:
The permeability barrier of nuclear pore complexes appears to operate via hydrophobic exclusion.
EMBO Journal 21, 2664-2671 (2002).
6.
S. Frey, D. Görlich:
A saturated FG-Repeat hydrogel can reproduce the permeability properties of nuclear pore complexes.
Cell 130, 512-523 (2007).
7.
S. Frey, R. P. Richter, D. Görlich:
FG-rich Repeats of nuclear pore proteins form a three-dimensional meshwork with hydrogel-like properties.
 unterdrückt die Passage von „normalen“ (inerten) Makromolekülen (A), erlaubt jedoch eine praktisch ungehinderte Passage von Importinen und anderen Kerntransportrezeptoren (B). Importine binden ihre Fracht im Zytoplasma und entlassen sie, sobald sie auf RanGTP im Zellkern treffen. Der Importin-RanGTP-Komplex passiert die Permeabilitätsbarriere dann in entgegengesetzter Richtung, im Zytosol wird GTP hydrolysiert, Ran entlassen und das Importin kann erneut Fracht laden. Das Zusammenspiel mit dem RanGTPase-System macht das Importin zu einer Proteinpumpe. Die eigentliche Passage durch die Permeabilitätsbarriere ist jedoch ein reversibler und energieunabhängiger Prozess.")
 erinnern. Ein solches FG-Hydrogel wurde in A auf eine gemusterte Unterlage platziert und photographiert. FG-Repeats sind hydrophile, lineare Polymere, die in regelmäßigen Abständen hydrophobe Flecken (FG-Motive) enthalten. Die Hydrogel-Bildung beruht auf einer Quervernetzung zwischen einzelnen FG-Repeat-Domänen, die u. a. durch hydrophobe Interaktionen zwischen den FG-Motiven (blaue Punkte) vermittelt werden (siehe B). Das FG-Hydrogel kann daher als dreidimensionales Sieb betrachtet werden, das kleine Moleküle ungehindert passieren lässt, aber den Durchtritt von inerten Makromolekülen (ocker) unterdrückt (siehe C). Dabei bestimmt die Maschenweite die Ausschlussgrenze (≈ 5 nm Durchmesser).")
, die Kernporen mithilfe eines Kern-Transportrezeptors (z.B. Importin) zu passieren. Die Größe solcher Transportkomplexe übersteigt die Maschenweite des FG-Hydrogels; sie müssen deshalb für ihren Durchtritt die im Wege liegenden Maschen des FG-Hydrogels kurzzeitig aufbrechen. Die Schemata A-D illustrieren diesen Prozess. Eine Masche wird durch Interaktion zwischen FG-Motiven (blau) gebildet und stabilisiert. Importine binden diese Motive, öffnen dadurch eine Masche und können das Hindernis samt ihrer Fracht passieren. Schemata E-F zeigen eine Kernpore mit FG-Hydrogel-Barriere. Importine (Imp) können Maschen lokal öffnen und sich im Hydrogel lösen. Sie werden dann gewissermaßen von der Permeabilitätsbarriere umflossen. Das stellt sicher, dass die Barriere intakt bleibt, selbst während große Objekte durch die Pore geschleust werden. Der im Gel gelöste Transportrezeptor kann die Barriere auf der anderen Seite wieder verlassen.")
. Schema des Messansatzes. Ein FG-Hydrogel wird so auf dem Objekttisch eines konfokalen Laser-Scanning-Mikroskops so positioniert, dass die Puffer/Gel-Grenze in der konfokalen Bildebene abgebildet wird. Der Einstrom zugegebener fluoreszenter Proteine in das Gel wird in Echtzeit mikroskopisch verfolgt. B) Aufnahme eines FG-Hydrogels, das mit einem inerten rot-fluoreszierenden Protein und einem grün-fluoreszierenden Importin-Fracht-Komplex inkubiert wurde. Das inerte Protein blieb ausgeschlossen, während der Importin-Komplex tief in das Gel eindrang. C)Durch konfokale Laser-Scanning-Mikroskopie wurden das far-red markierte FG-Hydrogel („Gel“) sowie der Einstrom eines rot-fluoreszierenden inerten Moleküls in dieses Gel visualisiert. Dieser Einstrom war sehr ineffizient, d.h. das Gel erwies sich als eine sehr effektive Barriere. D) Gleichzeitig wurde der Einstrom eines grün-fluoreszierenden Importin-Fracht-Komplexes verfolgt. Dieser Einstrom war so effizient, dass die Diffusion an die Gelgrenze zum geschwindigkeitsbestimmenden Schritt wurde. Der Komplex reicherte sich sehr stark im Gel an (Partitionskoeffizient > 100); die Aufnahmen sind daher im Gelbereich überexponiert. Entscheidend ist, dass der Importin-Fracht-Komplex nicht an der Puffer-Gel-Grenze stecken bleibt, sondern sich mit einer Geschwindigkeit in das Gel bewegt, die eine Kernporenpassage innerhalb von 10 Millisekunden gestatten würde. E) und F) Quantifizierungen der experimentellen Primärdaten von C und D. Dabei sind die Konzentrationsprofile der diffundierenden Moleküle zu verschiedenen Zeitpunkten ihres Einstroms in das FG-Hydrogel dargestellt.")